Reptiles
From Comparative Physiology of Vision
Remember to check the discussion tab at the top of the page.
| Reptiles | |
|---|---|
 |
|
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
Contents |
General Anatomy
Structure of the Eye
While birds and most reptiles have a ring of bones around each eye that supports the eyeball (the sclerotic ring), crocodilians lack these bones, just like mammals and snakes.
Some lizards and tuatara contain a third eye used for detecting changes in ambient light conditions, called a parietal eye, located on the top of their head. This eye contains a cornea, lens, and retina like the lateral eyes, but it is simpler in structure. Only two types of neurons are in this eye: photoreceptors and ganglion cells.
Eyelid
Snakes do not possess eyelids. A scale that is actually part of the skin, a spectacle, protects the eyelids. During ecdysis, the spectacle sheds from the eye along with the rest of its skin. Spectacles turn a light blue as it prepares to shed. If the spectacle is not removed during ecdysis, this can lead to eye damage.[1]


In crocodiles, a third transparent eyelid moves sideways across the eye (nictitating membrane). This eyelid protects the otherwise ‘open’ eyes when they submerge and attack under water. The third eyelid also conceals the liquid excretions produced by the crocodilian tear glands. However, if it is out of the water for any length of time, the resulting dryness in its eyes will cause visible tears to be formed. This is possibly why both factual and fictional accounts tell of crocodilians weeping over their prey on a river bank. They cannot focus underwater, meaning other senses are more important when submerged underwater.
Pupil
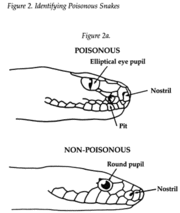
Snakes possess two different types of pupils, round and elliptical. Not always but usually, non-poisonous snakes have round pupils while poisonous snakes have elliptical (egg-shaped or cat-like). [2]
Snakes are unlike most mammals who change the shape of their lens to focus light on the retina. Most snakes do not possess these ciliary muscles and therefore cannot focus on stationary objects. [3]
A crocodilian’s pupil can close to a narrow, slit-like aperture in bright light, while opening to a full, circular pupil to allow maximum light collection in the dark. This is known as a stenopeic pupil, common among certain lizard species (i.e. the Tokay Gecko). So, they can hunt both during the day and at night, though they tend to hunt mostly at night and in the dim light of dawn and dusk. Bright sunlight means that it is time for basking on the river bank.
Retina
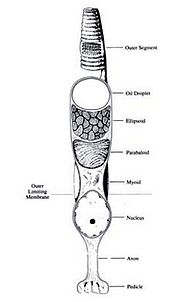
The retina is composed of two segements, the outer segement and the inner segement. The outer segment contains disks with photosensitive pigment molecules. The orientation and size of these disks determines whether the photoreceptor cell is a rod or a cone. The outer segment connects to the inner segment by a ciliary stalk. The inner segment may have any or all of the following: an oil droplet, an ellipsoid, a paraboloid, and a myoid. A nucleus lies beneath the region of the myoid and is connected to a synaptic pedicle via an axon. The synaptic pedicle communicates with other retinal cells.
The difference and sensitivity of each cells is dependent upon the differences in the outer and inner segments. Disk orientation of the outer segments determines whether a cell is a cone or rod whereas the amount of oil, oil fluorescence, and cell size and shape all have a unique impact on cell behavior. Similar to other non-mammalian vertebrates, some visual cells may be fused. Two identical photoreceptors that are fused, or paired, in which the two members are completely identical. Double photoreceptors have two dissimilar cells where one acts dominant and the other as an accessory. These cells are bound together and appear to be electrically coupled. [4]
Outer Segment
Like other vertebrates, the outer segment is composed of stacked disks within a plasma membrane. If the disks are discontinuous with the plasma membrane, they are a rod. However, if the disks are bulging against the membrane, they classified as a cone. Rods are responsible for vision at low illumination levels while cones are used for color vision at higher light levels.
The majority of photoreceptors in diurnal reptiles are cones. There some regions of the retina that exclusively contain cones. However, nocturnal reptiles experience a much greater quantity of rods than diurnal species. Comparatively, nocturnal reptiles have a similiar rod to cone ratios to that of humans.
Outer segments vary across species in size (length and diameter) and shape. The type of outer segment is dependent on behavior and habitat. The retinas of diurnal species have photoreceptors with narrow, short outer segments whereas nocturnal species increase light absorption by having long, thick, cylindrical outer segments. Nocturnal species that depend on vision for foraging have especially large outer segments.
In the parietal eye of lizards, the outer segment of the photoreceptor is oriented towards light entering the eye. This is opposite to the orientation of rod and cone cells, which have their outer segments embedded in the retinal epithelium.
Inner Segment
The inner segment organization is highly variable in reptiles. The ellipsoid is the only component present in all species of reptiles whereas snakes are the only reptile to not have an oil droplet as a part of their photoreceptors.
Oil Droplet
Oil droplets are located at the tip of the inner segment. Its primary function is to concentrate incoming light and focus it onto the outer segment. The oil droplet is present in all species of reptiles excluding snakes and some nocturnal lizards. Oil droplets are a feature of cone cells in reptiles (rod cells do not contain oil droplets).
Oil droplets contribute to light collection because of their high refractive indices. Incoming light is concentrated onto the base of the outer membrane, almost directly above the oil droplet. The focusing of light onto the membrane causes a change in membrane potential. Increased light sensitivity due to oil droplets can improve retinal sensitivity and increase contrast differentiation and motion detection.
Most oil droplets are yellow, orange or red and are a derivative of carotenoid pigments and their concentrations. The droplets act as a filter, only redirecting light of a certain wavelength range onto the base of the outer segment. For example, red sensitive cones with a peak sensitivity of 620 nanometers would have a red oil droplet that only focuses light onto the outer segment that has a wavelength greater than 550 nanometers. The relationship causes specific photopigments to always be paired with their optimal oil droplet.
Ellipsoids
Ellipsoids are found in every reptile photoreceptor cell. It is composed of mitochondria and refracts light, contributing to light-redirecting properties of the inner segment. In the case of snakes whom lack oil droplets, the ellipsoid has lipid packets in its membrane that redirect incoming light onto the base outer segment. The ellipsoid is responsible for providing energy rich phosphates to the outer segment. [5]
The ellipsoids of reptiles consists of densely packed mitochondria. The arrangement of the mitochondria differs between species, however. Alligators are have long mitochondria that are arranged parallel to the photoreceptor, similar to mammals. Diurnal lizards have a central mitochondrion that is larger than the surrounding mitochondria. The size and internal structure of the ellipsoid varies greatly between species.
Unique Visual Optics
Snakes use a combination of infrared vision (developed in the trigeminal nerve), variable (by species) visual acuity and color detection, limited eye mobility, and chemosensation to find prey and recognize features in their environment (including their keepers).
As with other reptiles, snakes have a yellow filter that absorbs ultraviolet light and protects the eyes. [6] [7]

Crocodiles see well during the day and may even have color vision, plus the eyes have a vertical, cat-like pupil that also gives them excellent night vision. The iris is silvery (a layer of crystals behind the retina at the back of the eye, present in many animals. This layer is known as the tapetum lucidum, and the crystals reflect the light that passes through the retina back onto the retina, thus increasing their ability to see in weak light), making their eyes glow in the dark.
Photo Transduction
The transduction of the photoreceptor of the parietal eye in lizards is unique to any vertebrate. In response to light, rod and cone cells decrease the amount of secondary signaling molecule, cGMP, inside the cell. This causes cation channels to close and the cell to hyperpolarize decreasing the rate in which the cell fires. The opposite is true for the photoreceptor of the parietal eye. Going from dark to light conditions causes a cascade of cGMP inside the cell which opens non-selective cation channels and allows calcium and sodium to enter the cell and depolarize it. The result is an increase in firing of the receptor cell. 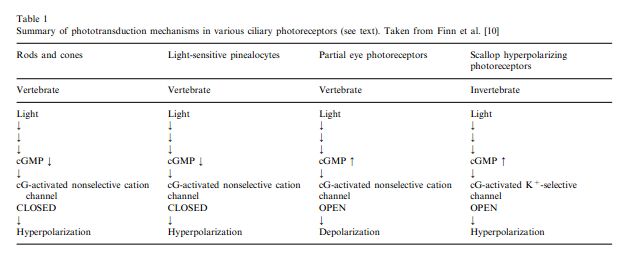
Color Vision
Lizard (including geckos) and turtle retinas contain multicolored oil droplets in their photoreceptors, so they can perceive color. The opsin proteins in the cones in the eye are "calibrated" to detect different wavelengths. In many species, this enables them to see into the higher wavelengths beyond the scope of unaided human vision: into the UV range. [6] [7]
The light receptors in crocodilians’ eyes include both cones and numerous rods, so it is assumed all crocodilians can see colors.
Motion Detection
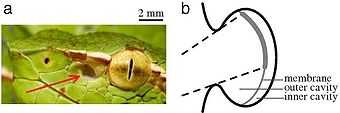
Pit vipers and boids possess infrared vision, or heat-vision, which allow them to effectively catch their prey. The detection system of their infrared vision consists of pit holes (not to be confused with the nostrils). Pit vipers possess one pit organ on either side of the head while pythons and boas possess several on each side of the head. The pit holes become activated and send signals through the nervous system to the brain. The brain then processes the information and reconstructs an image. [8] [9]
Neuronal Processing
The pit holes are composed of a heat-sensitive membrane which detect radiation in the form of infrared rays. This membranes contains sensory cells with special receptors that can detect the difference in heat between moving prey and its surroundings in milliKelvins. These receptors are known as TRPA1 channels.[10] The sensory cells are actually the terminal end of the trigeminal nerve. That is to say that the receptors are attached to the trigeminal nerve. Infrared waves are detected by these receptors located on the terminal nerve mass. In pit vipers, there are about 1,600 sensory cells per membrane. These proteins are temperature-sensitive ion channels that depolarize when heated. It is estimated that the sensitivity of these nerve fibers are greater than 0.001 °C. These nerve fibers are constantly firing at a very low rate. An increase in thermal radiation results in a reach of threshold. Neuronal firing increases as heat increases. These nerve fibers also possess an adaptive response. The pit organ will adapt to a repeated stimulus. If the stimulus is removed, the neuron will hyperpolarize and eventually reaches its resting rate. This period of adaptation lasts 50-150 msec.
In crocodilia there is also a small muscle (which is also seen in geckos) next to or upon the stapes, the stapedius, which probably functions in the same way as the mammalian stapedius muscle does, dampening strong vibrations. The upper and lower jaws are covered with sensory pits, seen as small, black speckles on the skin, the crocodilian of the lateral organ seen in fish and many amphibians. But they have a completely different origin. These pigmented nodules encase bundles of nerve fibers that respond to the slightest disturbance in surface water, detecting vibrations and small pressure changes in water, making it possible for them to detect prey, danger, and intruders even in total darkness. These sense organs are known as DPRs (Dermal Pressure Receptors). While alligators and caimans only have them on their jaws, they have similar organs on almost every scale on their body. The function of the DPRs on the jaws is clear, but it is still not quite clear what the organs on the rest of the body in crocodilians actually do. They are probably doing the same as the organs on their jaws, but it seems like they can do more than that, like assisting in chemical reception or even salinity detection
Higher Order Visual Perception
The "heat-sensing" proteins attached to the pit organ are innervated by the trigeminal nerve (Cranial nerve five). Information from the trigeminal nerve is relayed to the nucleus reticularus caloris in the medulla. Information the travels to the contralateral optic tectum. The optic tectum processes these infrared signals. Since point-like images are relayed to a membrane, the images appear blurry. However, these snakes possess edge detectors in the brain that turn a blurry image into a much clearer one. The edge detectors allow snakes to distinguish prey from its surrounding. [8] [9]
Evolutionary Significance
Recent studies have shown the parietal eye photoreceptor in lizards contains a mechanism for distinguishing colors. The cell contains two separate light detecting proteins, opsins; one that responds to blue light and another that responds to green light. The blue light sensing protein, pinopsin, decreases the amount of cGMP in the cell which hyperpolarizes the cell, while the green light sensing protein, parietopsin, increases the cGMP and causes depolarization and increased firing. Pinopsin is linked to the G-protein, gustducin, which is used in taste receptor cells and is much like the transducin G-protein found in rods and cones. The parietopsin is a yet to be classified protein, but it is apparently highly conserved in evolutionary history. It is linked to a G-protein called Go in these parietal photoreceptors and is reminiscent of a more ancient Go-protein pairing in scallops. The apparent age of the parietopsin-Go pairing compared to the more recently evolved pinopsin-gustducin/transducin pairing points to an evolutionary step towards dichromatic vision. Ancient photoreceptors contained a Go-mediated tranduction pathway and evolved a second gustducin/transducin-mediated pathway allowing antagonistic response to different wavelengths of light. This primitive form of color vision containing Go-mediated pathways was ultimately dropped leaving only the gustducin/transducin pathway that rod and cone cells use. Distinguishing colors in vision was moved to secondary neurons in the retina which provides more complexity.
References
- ↑ Douglas Mader, M.S., DVM, DABVP. Snake Anatomy. ReptileChannel.com http://www.reptilechannel.com/kid-corner/beyond-beginners/snake%20anatomy.aspx
- ↑ Pierce, Robert A. Snakes: Information for Missouri Homeowners. University of Missouri Extension. Missouri.edu. http://extension.missouri.edu/p/G9450
- ↑ http://www.icon.co.za/~mvdmerwe/anatomy.htm#eyes
- ↑ Carl Gans, 1992, University of Chicago, Sensorimotor Integration: Biology of Reptilia. http://books.google.com/books?id=Vkm1JpjiWr0C&pg=PA24&lpg=PA24&dq=reptile+photoreceptors&source=bl&ots=J6yC7TFiCt&sig=ODum4UjeSFNQqeNC8A3CLSLwDJ0&hl=en&ei=DbvSTorNN-jmiAKmidm7Dw&sa=X&oi=book_result&ct=result&resnum=4&ved=0CD8Q6AEwAw#v=onepage&q&f=true
- ↑ Michael BiernBaum, 1985, Department of Zoology, University of Wisconsin Frog Rod Outer Segments with Attached Inner Segment Ellipsoids as an In Vitro Model for Photoreceptors on the Retina. http://jgp.rupress.org/content/85/1/83.full.pdf
- ↑ 6.0 6.1 Sinclair, Sandra. 1985. How Animals See: Other Visions of Our World. Croom Helm, London.
- ↑ 7.0 7.1 Grace, Michael S. 1997, The visual system and non-visual photoreception. In: The Biology Husbandry, and Health Care of Reptiles. Lowell Ackerman, DVM, ed. Vol. I, pp. 325-341. TFH Publishing, Neptune City, NJ.
- ↑ 8.0 8.1 Zyga,Lisa. Snakes’ heat vision enables accurate attacks on prey. Physorg.com http://www.physorg.com/news76249412.html
- ↑ 9.0 9.1 http://rice.dreab.com/p-Pit_organ
- ↑ Agence France-Presse. Snakes 'see' with odd type of vision. The Science of Everything Cosmos. http://www.cosmosmagazine.com/news/3354/snakes-see-with-odd-type-vision?page=0%2C1
Allen & Unwin. 2006. Evolution’s Greatest Survivor Crocodile. Lynne Kelly’s Press. Australia.
Lisa Manson. June 2008. Husbandry Guidelines for The Freshwater Crocodile. Lisa Manson’s Press.
Burbidge A.A. (1987). The management of crocodiles in Western Australia. In: G.J.W. Webb, S.C. Manolis and P.J. Whitehead (Eds.), Wildlife Management: Crocodiles and Alligators. Surrey Beatty & Sons Pty. Ltd. in association with the Conservation Commission of the Northern Territory, Sydney
Finn, J. T., E. C. Solessio, and K.-W. Yau. "A CGMP-gated Cation Channel in Depolarizing Photoreceptors of the Lizard Parietal Eye." Nature 385.6619 (1997): 815-19.
Su, C.-Y. "Parietal-Eye Phototransduction Components and Their Potential Evolutionary Implications." Science 311.5767 (2006): 1617-621.
http://en.wikipedia.org/wiki/Crocodilia
Questions
What is the name of the "third eyelid" possessed by crocodilia and some lizards?
What is the purpose of the oil droplet in reptilian photoreceptors?
What is the name and function of the third eye in some lizards and tuatara?
