Spiders and scorpions
From Comparative Physiology of Vision
| Amphibians | |
|---|---|
 |
|
| Orb Weaving Spider | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Arachnida |
| Order: | Araneae |
Spiders and scorpions belong to the class Arachnida in the phylum Arthopoda. Arachnida is very diverse as a class, containing many constituent orders, including spiders, scorpions, ticks and harvestmen (known colloquially as daddy-long-legs, and not true spiders). Spiders and scorpions are generally predatory species, and employ various hunting and survival techniques which differ significantly based on the visual acuity of the species. Scorpions and most spiders, particularly orb-weaving species, rely much more on other senses than vision, such as chemo- and mechano-senses for catching prey. Hunting spiders, including wolf and jumping spiders, depend much more on vision and thus have more sophisticated methods of photo-reception than scorpions and orb-weaving spider varieties.[1][2]
Contents |
General Anatomy
Spiders and scorpions are characterized primarily by the presence of eight legs and two major body segments, the prosoma and the opisthosoma, also known as the cephalothorax and abdomen, respectively. It is in the prosoma where the extremities and sensory organs are located, including the eyes.

Spiders have eight or six eyes arranged on the prosoma, with two eyes in each of three or four rows across the carapace of the prosoma. These configurations are anterior median eyes, anterior lateral eyes, posterior median eyes, or posterior lateral eyes. The large, forward-facing eyes of a spider are the anterior median eyes, which have a very narrow field of view but excellent spatial resolution. The smaller forward-facing eyes are the anterior lateral eyes, which have a moderate field of view and moderate spatial resolution. The posterior eyes are more peripheral, secondary eyes, used for sensing motion with wide field of view. As such, the posterior eyes are extremely sensitive and tuned for nocturnal use.[3] Spiders with only six eyes do not have anterior median eyes[1]
Arachnids are the only class of Arthopoda to not have compound eyes, instead having camera-type eyes not unlike those of vertebrates[2].
Secondary eyes include posterior median and lateral eyes as well as anterior lateral eyes. Contrary to the anterior median eyes, the rhabdomes of secondary eyes point away from the light and lie proximally. Almost all secondary eyes possess a tapetum, which is a crystalline light reflecting layer that is absent from the main eyes. Eyes that possess a tapetum are divided morphologically into three major types, based on the shape of the tapetum. The three types of tapetum are the primitive type, which covers the entire back of the eye with only holes for nerves; the canoe-shaped tapetum, which resembles the primitive type but is split down the center; and the grated tapetum, which as its name implies is grated in shape, with many slits along it, resembling a sewer grate.
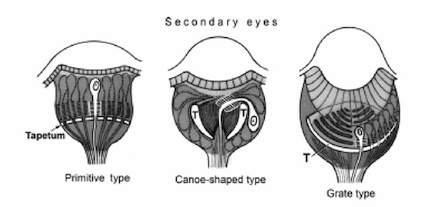
The tapetum of the secondary eyes lines the cup of the eye, so as to reflect light into backwards facing photoreceptor cells. In many secondary eyes, photoreceptors are exposed to both direct and reflected light, which increases sensitivity for low-light vision[1].
Unique Visual Optics
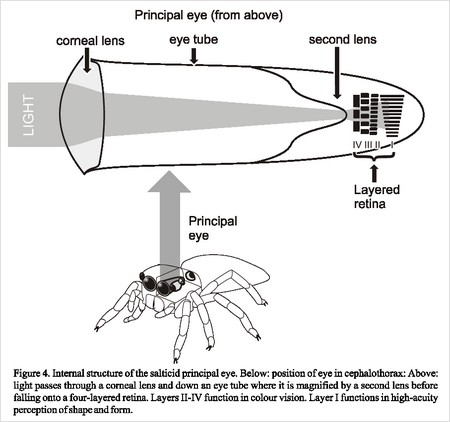
Eye structure in spiders can be classified into two types. The main eyes, which are always the anterior median eyes, generally maintain similar structure among different spiders. The second type, the secondary eyes, vary considerably among arachnid species and can further be divided into three subtypes.Unlike many eye structures, the anterior median eyes of spiders have two sets of lenses. The corneal lens, that is the lens near the outside of the eye tube, acts like a lens in any other eye. It changes shape in order to change the focal length based on what distance the object they are looking at is. The corneal lens has a long focal length to help traverse the long eye tube. At the end of the tube there is a second, concave lens. This acts like the lens in a telescope. That is the second lens magnifies the image which helps the spider see the shape and size of what they are looking at much better. This is especially helpful for hunting spiders, like the jumping spider, because it helps them identify prey. [4]
All spider eyes have light absorbing microvillar photoreceptors similar to other arthropods. These photoreceptors have the potential ability to sense polarized light. This special ability allows them to navigate by sensing the direction and angle of light rays from the sun. The light absorbing pigments are organized in fashion that responds optimally to light waves that vibrate parallel to the axis of the microvilli. The rhabdomere is the light absorbing structure and comprised of many microvilli which are arranged in parallel to each other through out the entire length of the rhabdomere. This is what causes the rhabdomere to respond optimally to lightwaves vibrating parallel to the microvilli. These rhabdomers are tuned into different angles of polerization based on thier arrangement in the retina. The light reflecting tapetum found the the majority of secondary eyes can enhance the sensitivity of polarized light perception as it acts as a polarizer as well.
Main Eyes
The structure and function of the anterior median eyes are especially unique in Salticidae (the jumping spider), and have long been known to be responsible for the jumping spiders excellent visual acuity. They can be distinguished by their abnormally large size, and appear to be black because of the absence of the light-reflecting tapetum. One challenge that Salticidae faces is that the lens is completely fixed within the carapace, significantly limiting the field of view for the spider. While initially seeming like a disadvantage, Salticidae compensates for its immobile lens in a couple of ways. The first and most practical way of dealing with this anatomical afflication is through the horizontal and lateral movements of the carapace. The structure and function of the Salticidae retina, however, is primarily responsible for the excellent vision that is characteristic of the jumping spider. The retinal structure is composed of 4 tiered layers of receptor cells, positioned in the central area of highest acuity. The first layer of receptor cells lies distally in relation to the cornea and is thought to be composed of a homogenous population of receptor cells that respond to green wavelengths. The retina lies within a 1mm eye cup and has come to be characterized as "boomerang" shaped. Each layer of these photo-receptors are sensitive to a specific wavelength of light. The first layer sits in the back of the retina, while the fourth layer is closest to the corneal lens. Layer 4 is suited to UV light, layers 2 and 3 to distinguishing color, and the first layer for spatial resolution.[5]
There is evidence that wolf spiders (Lycosidae) are able to navigate by polarized light using their primary eyes. When wolf spiders who lived on the shores of lakes were removed from their homes they would immediately return to their dens. But when these spiders were removed from their dens and placed on the opposite shore they would start to run out into the lake, suggesting they were using the polarization patters from the sunlight to navigate. When the experiment was repeated with the wolf spider's secondary eyes being covered the results were the same. However, when the primary eyes of the wolf spiders were covered they lost their ability to navigate at all. The structural and physiological basis for this polaization sensitivity in the primary, anterior medial eyes has yet to be discovered. [1] [6].
Secondary Eyes
The secondary eyes of most spiders have little to no spatial resolution, but very good sensitivity, much like rods outside the fovea of a human eye. Instead of providing the spider with imagery, they act as motion detectors. For some spiders without developed spatial resolution, this is probably as close as they come to "seeing" objects. In the case of the jumping spider, the secondary eyes serve to orient the spiders primary eyes towards moving objects. The anterior lateral eyes are somewhat unique in this context because they are intermediates, possessing the ability to "see" as well as effectively detect motion.
The secondary eyes of many spider species seem to be ideally developed for detecting light polarization. They have photoreceptors with consistently parallel microvili and the polarizing tapetum. In some species, the eye has even been shaped to give up acute spacial acuity in order to increase the receptive field and gain stimulus from larger regions of the sky. However, only one species, Drassodes cupreus, has demonstrated polarized light detection using its secondary eyes. A ground spider, Drassodes cupreus, leaves it nest every evening to search for prey. Using the polarization pattern produced by the sky it is able to successfully navigate back. When it's secondary eyes are covered with opaque, black paint. The spider looses its ability to navigate back home. The secondary eyes of Drassodes cupreus have a field of view that is able to receive stimulus from the entire sky. Each eye is arranged at opposing angles, so each eye responds optimally to different angles of light polarization. The tapetum of Drassodes cupreus is canoe-shaped, which is believed to be the favorable shaped tapetum for polarized light reflection. This canoe-shaped tapetum is found in many web-building spiders and it has been suggested that they are able to use their secondary eyes as compasses when constructing their webs. [7]
Color Vision
Orb-weaving spiders have not demonstrated much ability to distinguish color and likely posses only one photoreceptor pigment in all eyes (monochromatic). Some jumping spiders have been shown to possess trichromatic vision in the anterior median eyes, that is, three photoreceptor pigment types. Other jumping spiders seem to have only two different receptor pigmentations, but still maintain their ability to detect color. Green and UV spectra are the sensitivities found in most jumping spider eyes, while some species appear to have additional pigments in the UV/green or blue/green range.
The lateral eyes of hunting spiders, like the jumping spider, also only have a single curve of spectral sensitivity, while their anterior median eyes can have up to four different ones, making them tetrachromatic[8]. The unique thing about color vision in spiders is that, even with several different receptor types, some species seem to only have two spectral sensitivities. All of their photoreceptors have peak sensitivity in UV spectra (around 360 nm) and green spectra (520-540 nm), when examined individually. Normally, this would not allow the eye to detect colors. However, because of their layered retina, colored light is best focused on a different layer based on its wavelength. Even if peak sensitivities of each receptor are similar, they can function to determine more than two wavelengths based on their position in the layered retina. Such a system based on focal lengths has a major drawback in that some colors cannot be focused properly if they are too far away from the spider. Since multi-colored vision is only available for near objects, it's probably most useful to the spider in identifying color patterns on other spiders, such as potential mates. [5]
Motion Detection
Motion detection is a primary use of scorpion and spider eyes. Peripheral stimulation of the eyes by motion detection allows spiders and scorpions to orient themselves towards prey. The role of motion detection varies between species but is important to both orb-weaving and jumping or hunting spiders.
In jumping and hunting spiders, the field of view of the anterior median eyes (principle eyes) is less than one degree [3]. With such a narrow field of view, it is widely believed that the anterior median eyes are not sufficient for detecting moving objects. [9][3] However, the other eyes of the spider (secondary eyes) have a much wider field of view, which enables them to act as motion detectors that allow the spider to orient its principle eyes toward objects of interest. Currently, the exact functions of each pair of secondary eyes is not certain, but recent research is proving some of the answers. The anterior lateral eyes have a field of view between -25 and 60 degrees and have some spatial acuity. In 2010, Zurek demonstrated that in the species Servaea vestita, the anterior lateral eyes were able to detect prey even with the anterior median eyes were blindfolded[3]. This suggests that anterior lateral eyes have a more significant role in spatial detection than previously thought. The posterior eyes do not have any helpful spatial acuity and function only to orient the spider by detecting motion (in addition to their navigational and circadian functions).
The system of motion and shape detection is used extensively when a hunting spider is looking to capture prey. Because of the small field of view of the anterior median eyes the hunting spider will first see its prey in one or more of its secondary eye pairs. Once a prey is sighted the spider will turn its body to get the object into its field of view of its anterior median eyes and will continue to move as the prey does to keep it in the field. While the spider can detect movement and see an object up to 3 meters away it can only start the recognition process at about 30 cm. At this point the anterior median eyes begin working on the recognition process. During this process the muscles in the eyes are constantly moving the retina to scan the image, which is projected onto the central region of the retina, the fovea. If the prey moves closer or father away during this then the retinas are moved to compensate for this as well [1].
Higher Order Visual Perception
The central nervous system of spiders is divided into two major ganglia, the supraesophageal ganglion and the subesophageal ganglion. The names of these ganglia denote their position with regards to the esophagus of the spider. While afferent neurons from the spiders legs converge on the subesophageal ganglion, the supraesophageal ganglion connects to afferent neurons from all eight eyes. Visual neurons from the eyes synapse in the lobus opticus in the dorsal supraesophageal ganglion, into specific compartmentalized regions for each eye. Incoming nerves from each eye pass through three optic neuropils(regions containing a large amount of neuron axons and dendrites). These optic neuropils are not equal for each eye, however. The third and final neuropil in the pathway from the secondary eyes(posterior lateral, posterior median and anterior lateral) are small and located close to the front of the brain. The pathway from the main eyes(anterior median), is similar to that of secondary eyes, but terminates in a much larger neuropil in the central body of the superesophageal ganglion. The central body of the supraesophageal ganglion serves as an integration center for visual information and serves as the brain of the spider. Thus, visual information of the more perceptive main eyes can be used to coordinate the spider's movements and behavior, while secondary eyes serve auxilarly functions, such as detection of motion or polarized light [1]
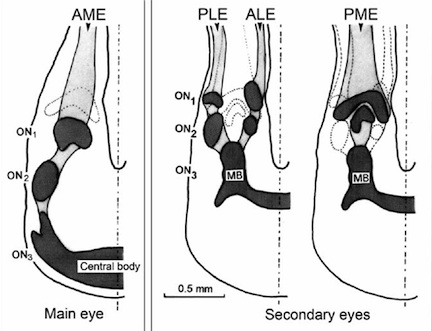
Photoreceptors in the eyes of spiders synapse on first order neurons, which then converge on second order neurons in the second optic neuropil, which is analogous to the medulla in vertebrate brains. These second order neurons then synapse on the third optic neuropil. [10]
In all spiders, the third optic neuropil occupies the same proportionate space in the brain, regardless of the size of dependence upon vision of the spider. The size and shape of this third neuropil is conserved among arachnids and their distant relatives, the horseshoe crabs. The number of lobes in the third optic neuropil varies among arachnids, with spiders possessing three lobes, and scorpions having four lobes. The retinotopic organization of the third optic neuropil is columnar, with different columns corresponding to the input from different eyes. Homologies with the optic neuropils of arthropods are also evident in the structure of the third optic neuropil of arachnids, including many cross hemispheric connections which are most developed in arachnids and horseshoe crabs.[10]
Scorpions
Scorpions behave, in many ways, similar to spiders in terms of vision. However, there are some differences worth noting. Like spiders, scorpions also have anterior median eyes that have a narrow field of view and pairs of lateral eyes that have more light sensitivity and quicker response. However, unlike spiders, scorpions can have up to five lateral eyes on each side, depending on the species, and they are not always paired up evenly. [11]
In many ways the anterior and lateral eyes are like the eyes of hunting spiders, like the jumping spider. However, there is an important difference in the lateral eyes. In the jumping spider the lateral eyes only have a single range of spectral sensitivity, with a maximum of 535-540 nm.[8] Scorpions' lateral eyes, on the other hand, have two different spectral sensitivities. One in the UV spectrum, peaking at about 371 nm., and the other a blue-green cone with a smaller peak somewhere in between 490 and 520 nm.[11]
Perhaps one of the most interesting difference between spider and scorpion photoreception is in the metasoma, or tail, of the scorpion. Research has shown that scorpions possess a special type of extraocular of photoreceptor in the metasoma of different types of scorpions. This is not a shared trait in all scorpions, but it is found in a variety of species. In addition, different types of scorpions will have different sensitivities in different parts of their metasomas. In otherwords, even in species that share the same spectral sensitivity in their metasoma, like the Heterometrus which has a sensitivity at 440 nm, they will not share what part of their metasoma is most sensitive to that light.[12] While the photoreceptors have been identified in these scorpions it is not completely understood what function they provide.[11]
Evolutionary Significance
(Some bullet points for expansion -Andrew)
-The development of both tetrachromatic eyes and monochromatic eyes in spiders indicates their visual system has at least two separate ancestors. The anterior median eyes may have developed from an ancestral anthropod's compound lenses
-The spectra of color vision in orb-weaving spiders is similar to honey bees, possibly to allow the spider to locate itself in areas attractive to honey bees.
-Polarized light detection provides navigation capability to spiders at night, even without any visual resolution.
-UV receptors in hunting spiders may help them find the webs of other spiders (which often refect UV light) they can eat
-Color and motion detection are also important in mating for some spiders. Jumping spiders exhibit bring colors and courtship dances, which have co-evolved with their visual abilities.
Questions
1. What are the physiological and functional difference between the primary (anterior median) and secondary (posterior median and lateral) eyes of spiders?
2. What evidence is there that Wolf Spiders use polarized light to navigate?
3. What unique characteristics do scorpions have in terms of vision?
References
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 1.6 1.7 Foelix, Rainer.(2010) Biology of Spiders, Oxford University Press.
- ↑ 2.0 2.1 Clemente, Christofer J., Kellie A. McMaster, Elizabeth Fox, Lisa Meldrum, Tom Stewart, and Barbara York Main. (2010)"The Visual System of the Australian Wolf Spider Lycosa Leuckartii (Araneae: Lycosidae): Visual Acuity and the Functional Role of the Eyes." Journal of Arachnology
- ↑ 3.0 3.1 3.2 3.3 Daniel B. Zurek, Alan J. Taylor, Christopher S. Evans, Ximena J. Nelson. (2010) "The role of the anterior lateral eyes in the vision-based behaviour of jumping spiders" The Journal of Experimental Biology, 213
- ↑ Harland, Duane P., and Robert R. Jackson. (2000)"'Eight-legged cats' and how they see - a review of recent research on jumping spiders (Araneae: Salticidae)" Cimbebasia 16: 231-240 PDF
- ↑ 5.0 5.1 Blest, A.D., R.C. Hardie, P. McIntyre, and D.S. Williams (1981)"'The Spectral Sensitivities of Identified Receptors and the Function of Retinal Tiering in the Principal Eyes of a Jumping Spider"
- ↑ Marshall, Justin. (1999). "Visual function: How spiders find the right rock to crawl under", Current Biology 9:R918-R921. PDF.
- ↑ M. Dacke, D.-E. Nilsson, E. J. Warrant, A. D. Blest, M. F. Land & D. C. O'Carroll. (1999). "Built-in polarizers form part of a compass organ in spiders". Nature. 401, pp. 470-473. PDF.
- ↑ 8.0 8.1 Yamashita, Shigeki and Hideki Tateda. (1976)"Spectral sensitivities of jumping spider eyes", Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology
- ↑ Daniela Neuhofer, Rudi Machan and Axel Schmid. (2009) "Visual perception of motion in a hunting spider" The Journal of Experimental Biology, 212
- ↑ 10.0 10.1 Homberg, Uwe. (2008). ‘’Evolution of the central complex in the arthropod brain with respect to the visual system.’’ Arthropod Structure and Development. 37. pp. 347-362.
- ↑ 11.0 11.1 11.2 Polis, Gary A. (1990) "The Biology of Scorpions", Standford University Press
- ↑ Rao, Geethabali and Kandula P. Rao. (1973) "A metasomatic neural photoreceptor in the scorpion", The Journal of Experimental Biology
