Cephalopods
From Comparative Physiology of Vision

Squid, octopods, cuttlefish and nautiluses are all members of the class cephalopoda, meaning "head-footed." This term stems from the way a cephalopod's body is constructed. The "feet" (usually called arms or mistakenly called tentacles) are attached to the part of the body containing the eyes (the "head") while the rest of the body is out in front of the head. Thus, the body does not connect directly to the arms.
Although we usually think of mollusks as benthic (bottom dwelling), the cephalopods have taken to a nektonic (swimming), rather than benthic, existence in the ocean.
Cephalopods share many characteristic molluscan traits with the rest of the creatures in the phylum. These include the presence of a mantle and mantle cavity, a radula in the mouth and a U-shaped digestive tract (useful for a creature in a conical shell, but not necessary for a cephalopod). Perhaps the most obvious difference between most cephalopods and other mollusks is the apparent lack of a shell. The octopods do not have shells at all, and the squid have a small chitinized internal shell. Nautiluses are the only cephalopods with an external shell. Nautiluses are found in the South Pacific and Indian oceans.
Cephalopods have the most well developed nervous systems of all molluscs, as well as the most well developed eye. Cephalopods rely more on vision than any other mollusc group.[1] Their eyesight is well suited for finding prey, avoiding predators and communication.
Comparative Anatomy
Cephalopods are known to have excellent senses and of these senses, their vision is perhaps the best studied. At a first glance cephalopod eyes look very similar to those of humans, whales and fishes. With the exception of the externally shelled and primitive nautilus, all cephalopods can perceive focused images, just like we can.
Cephalopods can see plane polarized light and have a district visual field. Humans have one focal point whereas cephalopods have many focal points. The advantage to many focal points is that his allows them to see movement very well . Cephalopods can not only see polarized light, they can also adapt this polarized light to their skin and change their skin to different colors. This implies an advantage of polarized light to underwater living.
The biological action for this is just now being understood. In a recent study, high concentrations of D-Asp (D-aspartate) was found in the retina of the cephalopods Sepia officinalis, Loligo vulgaris and Octopus vulgaris. D-Asp increases in concentration in the retina and optic lobes as the animal develops. In neonatal S. officinalis, the concentration of D-Asp in the retina is 1.8±0.2 μmol/g of tissue, and in the optic lobes it is 5.5±0.4 μmol/g of tissue. In adult animals, D-Asp is found at a concentration of 3.5±0.4 μmol/g in retina and 16.2±1.5 μmol/g in optic lobes (1.9-fold increase in the retina, and 2.9-fold increase in the optic lobes). In the retina and optic lobes of S. officinalis, the concentration of D-Asp, L-Asp (L-aspartate) and L-Glu (L-glutamate) is significantly influenced by the light/dark environment. In adult animals left in the dark, these three amino acids fall significantly in concentration in both retina (approx. 25% less) and optic lobes (approx. 20% less) compared with the control animals (animals left in a diurnal/nocturnal physiological cycle). The reduction in concentration is in all cases statistically significant (P=0.01–0.05). Experiments conducted in S. officinalis by using D-[2,3-3H]Asp have shown that D-Asp is synthesized in the optic lobes and is then transported actively into the retina. D-aspartate racemase, an enzyme which converts L-Asp into D-Asp, is also present in these tissues, and it is significantly decreased in concentration in animals left for 5 days in the dark compared with control animals. The dicarboxylic amino acids, D-Asp, L-Asp and L-Glu, may play important roles in vision.[2]
General Anatomy
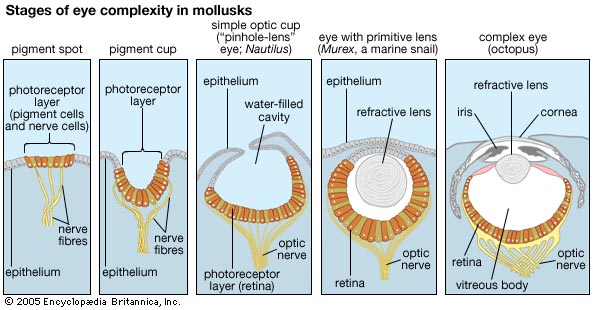
The cephalopod retina is everted in such a way that the distal end of photoreceptors lies directly behind the lens and points toward incoming light. As a result photoreceptor cells connect to the optic nerve behind the retina and, unlike in humans, there is no blind spot in the cephalopod eye.[1] Since there is very little difference in the refraction index of water and the eye a cornea is much less important in aquatic creatures than terrestrial. Of the cephalopods that have a lens, it is inflexible and roundish and made of crystallin. [3]
There are two eye types found in cephalopods: the pinhole type eye and the camera-type eye.
Pinhole Eye
Among cephalopods the pinhole type eye is found only in Nautilidiae, the pearly nautiluses. This eye type is unusual in that is open to the environment and is in direct contact with the water. It lacks a cornea or a lens to focus light and instead the eye functions like a pinhole camera. The iris forms a small adjustable opening, the pupil. Light passes through the pupil and forms an inverted image on the retina at the back of the eye. [1] The small size of the opening makes for a very dim image. Consequently, there is a trade-off between image sharpness and brightness with this eye structure. Since the nautilus doesn't have a lens, expanding the diameter of the pupil too much makes for a blurry image because too many photoreceptors are activated simultaneously.
Camera-type Eye
The coleoid cephalopods (which consists of all other living cephalopods) possess camera-type eyes similar to those of vertebrates. These eyes contain an iris, a rectangular pupil, a nearly circular lens, vitreous cavity and rhabdomeric photoreceptors forming a retina. [1]
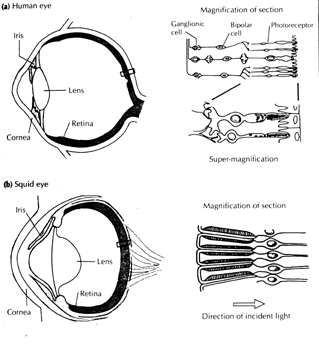
The pupil for different coleoid cephalopods varies. The octopus has a rectangular pupil, the cuttlefish a 'W' shaped pupil and the squid a circular pupil. They use their pupils to control the amount of light that hits the retina. The cephalopod eye resembles the vertebrate eye but is less sophisticated. Of all the cephalopods, the octopus has the most developed eye with a completely closed cornea. The octopus focuses by moving its entire (stiff) lens back and forth within the eye.[4]
Considering that cephalopods are water-dwelling creatures, one of their most interesting features is their ability to keep their eye orientation the same in relation to gravity regardless of their body position in the three dimensional water column. Essentially, their pupil will stay horizontally aligned regardless of the angle of their body. In order for this to work, they have a balance organ called a statocyst that controls the optic muscles. A statocyst is a sense organ that is half filled with both statoconia and endolymph and lined with hair cells. The statocyst gives the cephalopod input about orientation and acceleration similar to the utricle and saccule in human ears. [5] [6]
Unique Visual Optics
Visual pigments, the molecules in photoreceptors that initiate the process of vision, are inherently dichroic, differentially absorbing light according to its axis of polarization. Cephalopods have taken advantage of this property to build receptor systems capable of analyzing the polarization of incoming light. Polarized light is abundant in natural settings (often produced by scattering or reflection). Such polarization sensitivity has long been associated with behavioral tasks like orientation and navigation. However, only recently have we become aware that it can be incorporated into a high-level visual perception akin to color vision, permitting segmentation of a viewed scene into regions that differ in their polarization. By analogy to color vision, we call this capacity polarization vision. It is apparently used for tasks like those that color vision specializes in: contrast enhancement, camouflage breaking, object recognition, and signal detection and discrimination. While color is very useful in terrestrial or shallow water environments, it is an unreliable cue deeper in water due to the spectral modification of light as it travels through water of various depths or of varying optical quality. Here, polarization vision has special utility and consequently has evolved in numerous marine species, as well as at least one terrestrial animal. [7]
Photo Transduction
Many cephalopods have extraretinal photoreceptor cells called ‘epistellar bodies’ that allow researchers to collect information. The photoreceptor cells located in their retina are extremely difficult to use because they are long and thin and cannot easily be studied via intracellular recordings or patch-clamp work. [8] In a study conducted with the octopus Eledone cirrhosa, researchers traced the ionic mechanisms of phototransductions. The results concluded that extraocular photoreceptor cells depolarize by conducting sodium across the cell membrane in response to an increase in illumination. They used different ions as well as the pufferfish toxin tetrodotoxin (TTX) to determine that the response is diminished when sodium ion entry into the cell was reduced. [8]
Color Vision
Cephalopods are largely color blind. The research team Hanlon and Messenger trained octopuses to pick up black over white objects but could not train them to pick different colors. The visual pigments of over 20 species of cephalopods have been studied and with one exception all have been found to have only one visual pigment with a maximal absorption at 492 nm. The only cephalopod known to have color vision is the firefly squid Watasenia scintillans. This species of midwater squid is bioluminescent and has three visual pigments.[9]
Long have we associated photo receptors for polarized light with biological behavior and signaling and by nature all photo receptors are dichroic, meaning they absorb light rays with different polarizations in different amounts. Recent studies suggest, but have not proven, that cephalopods use polarized light akin to color vision. Polarized vision, or P-vision, can be used for things typically associated with color vision in humans; contrast enhancement, camouflage breaking, object recognition, and signal detection and discrimination. Cephalopods have long been associated with P-Vision this recent studys have shown color vision is not the only not needed but may have distinct disadvantage. The divergent evelosion of P-Vision and Color vision shows how nature can solve problems using divergent paths.[10]
Motion Detection
Neuronal Development and Processing

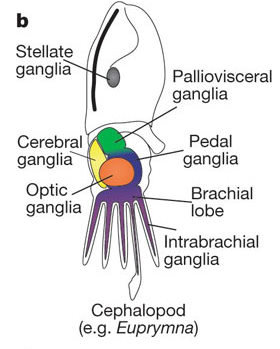
The development of the visual processing region of the cephalopod brain follows several specific developmental cues during embryogenesis. A study done by a lab at Georgia Tech found 3 distinct stages of neural development in cephalopods, specifically in the order Sepiolida [11]:
The first stage is referred to as “Accumulation of the ganglia.” This stage occurs after the differentiation of the eyes into two separate organs. Neuroblasts, or pre-neural stem cells, separate from the ectoderm and form four separate ganglial cell clusters. These clusters do not yet show any defined axon or dendrite structures, but will soon differentiate themselves into new structures.
The next stage sees the differentiation of the four ganglial clusters into three new masses (subesophageal, periesophageal, and supraesophageal), which further differentiate into four distinct "lobes": the cerebral, visceral, pedal, and optic lobe. This stage sees the appearance of dendrites and axons, called "neuropils," which are simply dense networks of axons, dendrites, and glial branches, first formed from the ganglia of the subesophageal and periesophageal masses. The subesophageal area was observed to grow quicker than it’s two counterparts. The suspected reasoning behind this is this area is used mainly for swimming and respiration, while the others are more for learning and other motor activity.
In the final stage of development, “growth of the neuropils” the lobes of the brain begin to form a more distinct shape of the cephalopod brain.
There is still more research to be done on the areas of neurotransmitters and genes. Pax6 is a gene that has been identified in the early stages of embryonic development. It is attributed to the development of the cerebral and optic ganglia, but it has not been researched enough to know the full functionality of this gene.
Biochemical Development
Cephalopods such as squids us a varied form of Rodopsin to see polarized light. D-Asp (D-aspartate) is an endogenous amino acid found in the nervous and endocrine system of various animals[2]. This amino acid is critical for polarized light detection. The common octopus was found to have this amino acid in the rhodopsin scientist have long studied the eye sight of Cephalopods and this amino acid is a common amino acid between human and cephalopods vision Cephalopods use this as a neurotransmitter directly to the photo luminescent skin and humans use it in high order brain functions. D asp is also found in large number of mammals and fish in high levels in vision cortex. The team of Salvatore D'Aniello,Patrizia Spinelli, Gabriele Ferrandino, Kevin Peterson, Mara Tsesarskia, George Fisher, and Antimo D'Aniello therize that it plays an important role to vision or visual processing. Table 1 D-Asp acid in the retina of adult S. officinalis, L. vulgaris and O. vulgaris Values are the means±S.D., expressed as μmol/g of tissue obtained from eight adult animals using the HPLC and the colorimetric methods. D-Asp* D-Glu+NMDA† Other D-amino acids‡ HPLC method α-Oxoacid method H2O2 method α-Oxoacid method H2O2 method S. officinalis (160–200 g body mass) 2.60±0.30 0.35±0.05 0.33±0.04 0.48±0.06 0.44±0.05 O. vulgaris (1200–1500 g body mass) 2.30±0.25 0.30±0.06 0.28±0.05 0.35±0.06 0.40±0.09 L. vulgaris (130–150 g body mass) 1.60±0.20 0.25±0.05 0.26±0.04 0.35±0.03 0.32±0.04 Taken from Biochem J. 2005 March 1; 386(Pt 2): 331–340. Published online 2005 February 22. Prepublished online 2004 October 18. doi: 10.1042/BJ2004107 This chart shows various methods style to determine mass of neurotransmitters per gram. D-amino acids have very important roles in vision. Rhodopsin uses it to form different polarized light receptors in Cephalopods. Conclusion of the study show that L-Glu and L-Asp and more importantly D-Asp is used the most but as the process that creates this amino acid is not known at this time further studys will be done to determine pathways.[2]
Phototransduction In Skin
There has been some evidence for light sensing capabilities in the skin of some cephalopods. Research has found opsin transcripts in fin and ventral skin samples of cuttlefish (Sepia officinalis). The corresponding amino acid sequence was identical in retinal and fin tissue samples but different from ventral tissue samples by one amino acid. Cuttlefish have been shown to be colorblind and thus color discrimination by skin opsins is unlikely as these opsins would be identical to those found in the retina. Chromatophores and iridophores, which produce changeable colors and patterns in the skin, could be involved in color discrimination. However, protein expression in cuttlefish has not yet been demonstrated and thus these dermal opsins may not be functionally significant. [12]
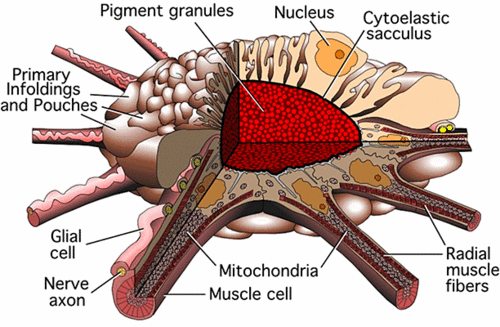
Chromatophore is a pigment-containing and light-reflecting cell, found in various invertebrate and cold-blooded vertebrate animals, that can help bring about changes in color or brightness in the organism. Among animals with such cells are various crustaceans, cephalopods, amphibians, fish, and reptiles. Through aggregation or dispersion of the pigment and reorientation of reflective plates in single cells (in most animals) or through the muscular movement of complex chromatophore organs (in cephalopods), the color or brightness of the entire organism or part of the organism can change, which is useful for camouflage, signaling, temperature regulation, and protection from radiation. Mature chromatophores are grouped into subclasses based on their color (more properly, "hue") under white light: Xanthophores (yellow), erythrophores (red), iridophores (reflective/iridescent), leucophores (white), melanophores (black/brown), and cyanophores (blue).
Chromatophores are largely responsible for generating skin and eye color in cold-blooded animals and are generated in the neural crest during embryonic development. In a cephalopod, like an octopus and cuttlefish, the complex chromatophore organs and neural control allow very rapid changes in colors, color patterns, and color intensity to allow the organism to blend instantaneously with the background. Unlike cold-blooded animals, mammals and birds have only one class of chromatophore-like cell type, the melanocyte. Although the term "chromatophore" also can refer to colored, membrane associated vesicles (organelles) found in some forms of photosynthetic bacteria. In addition to their obvious value for the animals with chromatophores, these cells are being explored for potential application for people, including modeling diseases, use in appetite suppression and tanning, and as biosensors and for rapid disease detection. These cells also add greatly to the diversity and wonder of nature for people. Cephalopods such as octopus have complex chromatophore organs controlled by muscles to achieve this. While Coleoid cephalopods have complex multicellular "organs" that they use to change color rapidly. This is most notable in brightly colored squid, cuttlefish, and octopuses.
Each chromatophore unit is composed of a single chromatophore cell and numerous muscle, nerve, sheath, and glial cells (Cloney and Florey 1968). Inside the chromatophore cell, pigment granules are enclosed in an elastic sac, called the cytoelastic sacculus. To change color, the animal distorts the sacculus form or size by muscular contraction, changing its translucency, reflectivity, or opacity. This differs from the mechanism used in fish, amphibians, and reptiles, in that the shape of the sacculus is being changed rather than a translocation of pigment vesicles within the cell. However, a similar effect is achieved.
Octopuses operate chromatophores in complex, wavelike chromatic displays, resulting in a variety of rapidly changing color schemes. The nerves that operate the chromatophores are thought to be positioned in the brain, in a similar order to the chromatophores they each control. This means the pattern of color change matches the pattern of neuronal activation. This may explain why, as the neurons are activated one after another, the color change occurs in waves (Demski 1992). Like chameleons, cephalopods use physiological color change for social interaction. They are also among the most skilled at background adaptation, having the ability to match the color, pattern, intensity, and the texture of their local environment with remarkable accuracy.[13]
Some oceanic squid have well-developed extra-ocular photoreceptors that measure downwelling light and are involved in adjusting ventral counterillumination. [12]
Higher Order Visual Perception


The study of the visual perceptive capacity of cephalopods remains speculative, yet promising. Studies have shown cuttlefish to be highly adept at orienting their bodies based of surrounding visual cues, such as directionally-oriented black & white patterns, a useful adaptation for proper camouflage in a variety of situations. [14] Though color blind, cuttlefish have been shown in studies to be adept at contrast resolution, able to differentiate between shades of gray down to 15% graded difference, comparable to humans' far greater contrast threshold of 2%. [15] It is unknown how color-blind cephalopods are able to camouflage themselves within hue-rich environments so well in the presence of di- and tri-chromatic predator species, suggesting greater capacities of visual processing than science can currently speculate.[16]
What is known is that cuttlefish' adaptive camouflage ability is proportional to the visual richness of their rearing environment. Cuttlefish raised uniform-grey environments showed greater disrupted capacity to camouflage in more contrasted environments later on than those raised in environments of high-contrast [17]

Evolutionary Significance
The likeness between the camera-type cephalopod eye and the vertebrate eye has been used as a classic example of convergent evolution for almost 140 years. Convergent evolution is the independent evolution of similar structural or functional components in two or more unrelated or distantly related lineages. The other possible explanation is parallel evolution wherein two lineages evolve from a common ancestor and in a common trajectory towards a similar endpoint, resulting in the similar or shared trait in different species. The convergence theory of the camera-type cephalopod eye is now being challenged. A conserved gene network has been discovered which includes the gene Pax6. Pax6 governs eye organogenesis in both vertebrate and invertebrate lineages. This gene conservation may indicate a common origin of eyes in vertebrates and invertebrates. If this is the case then the similarity between the camera-type cephalopod eye and the vertebrate eye would be a result of constraint in development and parallel evolution. However, it is both possible and probable that this likeness is the result of a combination of both convergent and parallel evolution, as the eye is a complex organ with many components which may evolve independently of one another. [1]
Cephalopods of Interest

The colossal squid Mesonychoteuthis hamiltoni has eyes of more than 27 cm across (about 11x that of human eye). [1]
The firefly squid Watasenia scintillans has color vision, with three retinal pigments, and can use ATP to fluoresce!
Exam Questions
1)What is P-Vision and why is it important to underwater organisms?
2)What unique connections do Cephalopods have between vision and skin?
3)What are Pinhole type and Camera type eyes?
Notes and References
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 Serb, Jeanne M. and Eernisse, Douglas J. Charting Evolution’s Trajectory: Using Molluscan Eye Diversity to Understand Parallel and Convergent Evolution. http://www.springerlink.com/content/1741pu336457n37u/fulltext.pdf
- ↑ 2.0 2.1 2.2 D'Aniello, Salvatore, et al. Cephalopod vision involves dicarboxylic amino acids: D-aspartate, L-aspartate and L-glutamate. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1134798/
- ↑ Map of Life - "Camera eyes of cephalopods." http://www.mapoflife.org/topics/topic_7_Camera-eyes-of-cephalopods/ December 02, 2011.
- ↑ Vendetti, Jann. The Cephalopoda: Squids, Octopuses, Nautilus and Ammonites. University of California Museum of Paleontology. 2006. http://www.ucmp.berkeley.edu/taxa/inverts/mollusca/cephalopoda.php
- ↑ http://www.ebiomedia.com/what-animal-has-a-more-sophisticated-eye-octopus-or-insect.html
- ↑ http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1692077/pdf/9415917.pdf
- ↑ Integrative and Comparative Biology Coverage: 2002-2005 (Vols. 42-45)(Vol. 46, No. 1 - Vol. 50, No. 6).
- ↑ 8.0 8.1 Cobb, Christopher S., and Williamson, Roddy. Ionic Mechanisms of Phototransduction in Photoreceptor Cells From the Epistellar Body of the Octopus Eledone Cirrhosa. http://jeb.biologists.org/content/202/8/977.full.pdf
- ↑ Mäthger, Lydia M., et al. Color blindness and contrast perception in cuttlefish (Sepia officinalis) determined by a visual sensorimotor assay. http://www.mbl.edu/news/features/pdf/mathger_etal_visres.pdf]
- ↑ Thomas W. Cronin, Nadav Shashar, Roy L. Caldwell, Justin Marshall, Alexander G. Cheroske1, Tsyr-Huei Chiou1 "Polarization Vision and Its Role in Biological Signaling ." Oxford Journals | Life Sciences | Integrative and Comparative Biology. N.p., n.d. Web. 26 Nov. 2011. <http://icb.oxfordjournals.org/content/43/4/549.full>
- ↑ Shigeno, S, K Tsuchiya, and S Segawa. “Embryonic and Paralarval Development of the Central Nervous System of the Loliginid Squid Sepioteuthis Lessoniana.”J Comp Neurol, 437.4 (2001): 449
- ↑ 12.0 12.1 Mäthger, Lydia M., et al. Evidence for distributed light sensing in the skin of cuttlefish, Sepia officinalis. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2936158/?tool=pmcentrez
- ↑ Cloney, R.A., and E. Florey. 1968. Ultrastructure of cephalopod chromatophore organs. Z Zellforsch Mikrosk Anat 89: 250–280. PMID 5700268
- ↑ Justine J. Allen, Lydia M. Mäthger, and Roger T. Hanlon "Cuttlefish use visual cues to determine arm postures for camouflage" Proc. R. Soc. B published online before print May 11, 2011, doi:10.1098/rspb.2011.0196 (http://rspb.royalsocietypublishing.org/content/early/2011/05/10/rspb.2011.0196.full)
- ↑ Lydia M. Mäthger, Alexandra Barbosa, Simon Miner, Roger T. Hanlon, Color blindness and contrast perception in cuttlefish (Sepia officinalis) determined by a visual sensorimotor assay, Vision Research, Volume 46, Issue 11, May 2006, Pages 1746-1753, ISSN 0042-6989, 10.1016/j.visres.2005.09.035. (http://www.sciencedirect.com/science/article/pii/S0042698905004943)
- ↑ Chiao CC, Wickiser JK, Allen JJ, Genter B, Hanlon RT "Hyperspectral imaging of cuttlefish camouflage indicates good color match in the eyes of fish predators." Proc Natl Acad Sci U S A. 2011 May 31;108(22):9148-53. Epub 2011 May 16. (http://www.ncbi.nlm.nih.gov/pubmed/21576487)
- ↑ Lee, Yi-Hsin; Yan, Hong Young; Chiao, Chuan-Chin "Visual contrast modulates maturation of camouflage body patterning in cuttlefish (Sepia pharaonis)" (http://psycnet.apa.org/?&fa=main.doiLanding&doi=10.1037/a0019461)
External Links
Cephalopod vision involves dicarboxylic amino acids: D-aspartate, L-aspartate and L-glutamate:
