Crustaceans
From Comparative Physiology of Vision
Vision in a Shell:
The Story of Crustacean Vision
| Lysmata debelius
(fire shrimp) |
|
|---|---|
 |
|
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Subphylum: | Crustacea |
| Class: | Malacostraca |
| Order: | Decapoda |
| Infraorder: | Caridea |
| Family: | Hippolytidae |
| Genus: | Lysmata |
| Species: | L. debelius |
| Binomial name | |
| Lysmata debelius Bruce, 1983 |
|
Contents |
General Anatomy
Crustaceans are part of the phylum arthropoda, a group of creatures that includes spiders, scorpions, insects, and mites. The crustaceans share the common features of the arthropods: they have an exoskeleton that provides support and protection and the body structure is built around pairs of jointed, segmented appendages. The eyes of these creatures follow this pattern and are found in pairs. The general structure of eyes varies from species to species; some are simple structures that do no more than detect the presence of light (such as the horseshoe crab) while others are complex structures that can receive multiple types of light including polarized and ultra violet light (such as the mantis shrimp). In two of the largest orders, the Isopoda and Amphipoda, there is no carapace and the eyes are sessile (immovable) rather than stalked.
Crustaceans occupy terrestrial, fresh- and saltwater habitats, and range in size from the Japanese spider crab (up to 3.8 meters and 19 kg) to the microscopic Stygotantulus stocki (0.1 mm). Some of the most well-known crustaceans, shrimp, crab and lobster, belong to Malacostraca, one of eight classes.
Multiple Photoreceptors
Crustaceans can have one or more types of photoreceptors[1] which include:
Compound Eyes
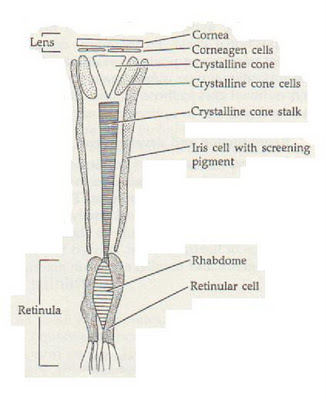
Many crustaceans have the type of eyes called compound eyes. Each eye is composed of many ommatidia. Each ommatidium is made of a cornea, a crystalline cone, and rhabdom and photoreceptor cells which are covered in pigment cells. The cone communicates with the rhabdom, a transparent tube in the center of the photoreceptors, which relays information to the optic nerve. Each rhabdom is paired with an individual optical system resulting in limited spatial resolution. All of the ommatidia focus to one point called the fovea. Some crustaceans have special adaptions for them to see in different ways. Mantis shrimp have specific rows of ommatidia to detect specific wavelengths of light.
While the compound eye has limited resolution relative to its size, it is able to provide possibilities unavailable to the more compact "camera eye" design. All of the photoreceptors in a single ommatidium are able to view an identical field, which can then be imaged onto several photoreceptor types allowing for spectral (color) or polarizational perception. Compound eyes have a very flexible geometry, so ommatidia in well-separated areas of the eye are able to view the same spatial location. This greatly multiplies the ability of parallel processing.
In mantis shrimp, much of the image analysis takes place early in the visual process which produces an analyzed, simplified stream of information flowing to higher visual centers.[2]
Nauplius Eyes
Nauplius eyes are usually present in larval stages, but can continue through adulthood. In clam shrimp, the nauplius eye consists of four cups of inverse sensory cells separated by a pigment layer and a tapetum layer. The pigment and tapetum layers contain two different kinds of pigment granules, the inner pigment layer consisting of large, dark colored granules, and the outer tapetum layer consisting of small, lightly colored, reflective pigment granules.[3]
Frontal Organs
In the absence of compound eyes, as in ostracoda and maxillopoda, frontal organs develop into complicated mirror, lens-mirror, and scanning eyes. Both nauplius eyes and frontal organs send their axons to the anterior region of the cerebrum.
Intracerebral Ocelli
A small simple eye, found in many invertebrates, usually residing in the brain as clusters of photoreceptors and a single lens.
Caudal Photoreceptors
Simple photoreceptors found in the sixth caudal ganglion.
Unique Visual Optics
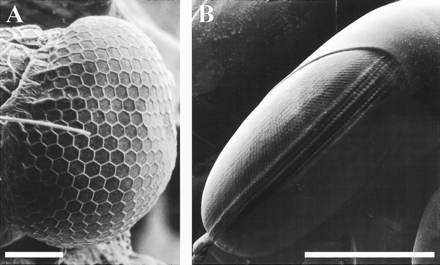
A. Compound eye of the first planktonic larval stage, anterior view. The eye is stalked and consists of a flattened spherical array of similar-sized ommatidia with neat hexagonal packing. Scale bar: 100 μm.
B. Lateral view of the compound eye of an adult. Note the ovoid shape of the eye and the very complex ommatidial array, including six raised rows of ommatidia that form the equatorial midband. Scale bar: 1 mm
Larval Vision
Larval vision is primarily focused on orientation in the water column, vertical migration, and avoidance of predators.[4] Euphausiid, decapod, and stomatopod larvae possess compound eyes. This creates a problem in planktonic crustacean larva as their passive defense is transparency and photoreceptors absorb light causing them to be less transparent. However, the larvae can condense their retinas to a tiny clump in the eye's center, packing the visual and screening pigments into a compact mass.[4] This is the apposition type because the eye permits the smallest retinal radius. Each photoreceptor unit in an apposition compound eye receives input through a single optical system.[4] The radially ordered photoreceptors are tightly grouped into a sphere in the center of the eye. The photoreceptors are seperated by screening pigments which lie next to and underneath the receptors to prevent light from entering in the wrong end. Larval eyes contain several types of pigments. Many of these are used to control the distribution of light in the retina or to reduce the retina's contrast against the background. They also have the photosensitive visual pigments. These visual pigments have been identified as typical rhodopsin with the middle wavelength absorption. Only one spectral class has been found in all the species. During their development, some crustaceans simply enlarge the larval eye, adding ommatidia as the animal grows. Others entirely discard the larval apparatus, literally pushing it aside and replacing it with a new optical and retinal array.[4]
Adult Vision
Adult vision also has to include navigation, prey recognition and capture, spatial vision, mate selection, and communication. The adult visual system is more complex than in larval stages and requires specialized ocular adaptations.[4]
Color and Polarized Vision

Compound eyes with visible mid-band of Odontodactylus scyllarus (Peacock mantis shrimp).
Color vision in crustaceans involves some of the same principles as it does in other forms of life but with distinct differences. One of the differences is that most crustaceans are aquatic. In order to see in color, they must have special pigments within their eyes that filter the light to compensate for the water that the light passes though. The pigments however, are similar to the pigments that other life forms use to see in color. Crustaceans, for the most part, do not require color vision to survive. However, color vision can confer a distinct advantage in some ecological niches; being able to tell what type of food source is available is important, and with color vision, this process may be easier. Color vision may also play a role in the selection or identification of a mate. In crowded coral reefs that may hold thousands of species of different organisms, the riot of color and the variety of shapes could present creatures with a problem of identification. Without color vision, the ability to differentiate between a potential mate and a potential meal could be difficult.
One example of color vision in crustaceans is the mantis shrimp. Mantis shrimp have color vision and are capable of processing ultra violet light, linear polarized light, and also circular polarized light. The general structure of a mantis shrimp eye is typical of a compound eye, with some subtle differences. Each eye is subdivided into three hemispheres. This separation into three sections gives each eye trinocular vision meaning that each eye is capable of depth perception. The upper hemisphere and the lower hemisphere contain ommatidia that are typical of crustaceans: they detect movement and are black and white vision only. Each eye also contains a mid-band, a group of ommatidia arranged in rows. These mid-band ommatidia are specialized for spectral and polarized vision.
The types of receptors present in the mid-band are vastly different from the hemispheres that the mid-band divides. This special section contains the more complex ommatidia including multiple types of pigmented cones. Mantis shrimp can have up to 12 different types of color processing pigments. Compared to a human's 3 different color pigment types, the shrimp must be capable of processing much more of the spectrum.
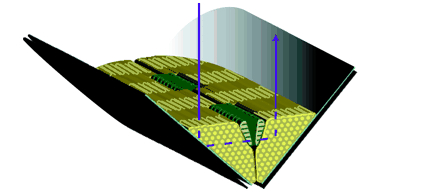
Because most dichroic visual pigments of arthropods are oriented in small, parallel, tubular membranes (mircovilli), they are naturally sensitive to the plane of incoming polarized light. Crustaceans take advantage of this sensitivity to polarized light by producing ommatidia that contain two photoreceptor classes. Each class has microvilli which are parallel but perpendicular to the microvilli of the other receptor class. This produces a system with two analyzers each of which is maximally sensitive at an e-vector orientation perpendicular to each other. In mantis shrimp, all ommatidium are polarization-sensitive.[2]
The eyes of mantis shrimp are situated on mobile stalks and move independently from one another granting these crustaceans a uniquely distinct and acute view of the world.
Light polarization is the direction in which light vibrates. This is different from the speed of light oscillation which is perceived as color. Many species, including the mantis shrimp, have the ability to see linear light polarization: the back and forth oscillation of light in a straight line after being reflected through water or off of a transparent membrane at different angles. The mantis shrimp also has the rare ability to see circular light polarization—the oscillation of light in a continuously circular direction. Circular light polarization occurs in two different directions, clockwise and counter-clockwise, both of which the mantis shrimp is capable of processing. It is speculated that the detection of circular polarized light is an attribute of reproduction due to the sex-specific reflection of circularly polarized light, although the same structures of the eyes are found in both male and female mantis shrimp. Other theories suggest that the sensitivity to circular light polarization is not a separate ability, but a component of the overall acuity of polarized light detection.
The ability to detect partially polarized light (the most common form of polarized light) gives an organism the heightened ability to detect surface texture or orientation of an object and is most commonly used by organisms for navigational purposes.
Evolutionary Significance

A horshoe crab (left) is considered a "living fossil", while a mantis shrimp (right) has evolved to have many specialized features.
Since eyes are present in all crustaceans, this universal presence demonstrates that eyes and vision have always been important for crustaceans throughout their evolutionary past. The evolution of comprehensive and complex visual systems may be attributed to the environment in which these creatures live. Ancestral malacostracans lived in the oceans some 200 million years before dinosaurs roamed the earth. Today, they constitute nearly 75% of all crustacean species and include lobsters, crabs, crayfish and shrimp. All of these different types have compound eyes, each developed to a different degree depending on selective pressures. Fossils dated around 500 million years ago show the slow evolution of crustacean eye structures, and give evidence of the first stalked compound eyes in existence. These fossils show that typical malacostracans had stalked eyes, 19 segments plus a telson (the posterior-most extension of the last body segment). Certain species have changed very little over evolutionary time this would indicate that there is very little selective pressure at work in the environment in which these types of creature live. For instance the horseshoe crab is refered to as a "living fossil" because it has changed very little or not at all in the course of evolutionary time. Selective pressures have had a great effect on other species. A prime example of the ability to adapt is the group of stomatopoda known as "mantis shrimp". The eyes of these creatures are vastly different than the eyes of the horseshoe crab. Clear evidence that the crustaceans are capable of adaptation and evolutionary modification. This ability to adapt is clearly demonstrated by the experiments of Cheroske, Cronin, and Caldwell, in which mantis shrimp were used in a controlled environment that manipulated the type and intensity of light exposure. The results seem to indicate that the light intensity is the more important factor: the shrimp can adapt to use the light available so long as it is provided in strong enough intensity. Evolution of crustacean eyes has been an ongoing process for millions of years. It is one of the few examples of evolution that we have fossil evidence for. Being able to refer to the fossil record provides a unique view of the process of evolution in crustaceans as well as a possible guide to other animals' evolutionary past.
References
- ↑ Meyer-Rochow, V. (2001). The Crustacean Eye: Dark/Light Adaptation, Polarization Sensitivity, Flicker Fusion Frequency, and Photoreceptor Damage. Zoological Science (VSP International Science Publishers), 18(9), 1175.
- ↑ 2.0 2.1 Cronin, Thomas W., & Marshall, Justin (2001) Parallel Processing and Image Analysis in the Eyes of Mantis Shrimp. Biology Bulletin. 200: 177-183
- ↑ Reimann, A., & Richter, S. (2007). The nauplius eye complex in “conchostracans”(Crustacea, Branchiopoda: Laevicaudata, Spinicaudata, Cyclestherida) and its phylogenetic implications. Arthropod structure development, 36(4), 408-419.
- ↑ 4.0 4.1 4.2 4.3 4.4 Cronin, Thomas W., & Jinks, Robert N. (2001) Ontogeny of Vision in Marine Crustaceans.
- ↑ http://jeb.biologists.org/content/204/14/2481/F6.expansion.html
http://jeb.biologists.org/content/206/2/373.full
http://www.plosone.org/article/info:doi/10.1371/journal.pone.0002190
http://www.science20.com/science_20/eye_evolution_gets_its_war_and_peace-84766
Questions for Final
1. Why do some crustaceans have color vision while others have black and white?
2. What is a “living fossil”? Give an example of a crustacean that is a "living fossil".
3. What is a unique ability of mantis shrimp vision that most other species do not have?
