Birds
From Comparative Physiology of Vision
Out of all its senses, birds rely tremendously on their vision. Since their reliance on vision is so significant, birds have developed unique characteristics in processing these types of information over 150 million years of natural selection. Avian eyes occupy a much larger volume of their head than do the eyes of other mammals. Large eyes are necessary in order for them to find food, identify potential mates, escape from predators, and aid in general orientation. Their remarkable color perception, spatial and temporal resolution, visual learning and memory capabilities have been diligently studied over the past 40 years.

| Avian | |
|---|---|
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Aves |
Contents |
General Anatomy
Lens and Cornea
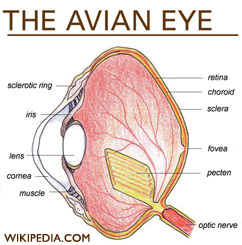
Birds eyes are proportionately much larger than human eyes. Eagles and owls have eyes of a similar size to humans, while ostrich eyes are nearly twice as large as the human eye. It is not uncommon among song birds for their eyes to be the same size as their brains.[1]
Birds eyes are not spherical. In fact, the exact shape differs depending on the species, unlike mammals.
The massive size of avian eyes allow for a sharp, large images, while the enlarged pupil allows more light to enter the eye, enabling birds to see in much dimmer light. [1]
The fovea is packed with 400,000 cones per square millimeter - twice that of a human. Hawks have even more, coming in at 1,000,000! The high density of retinal cells enables them to see in much finer detail then human. [2]
The outer most portion of the eye, the sclera, contains cartilage just as it does in reptiles. Its surface is opaque, turning clear in exposed regions, and it coordiantes with various other tissues of the eye to form the cornea. (STURKIE) The relative size of the sclera changes depending on the species, remaining very small in underwater swimming birds and large in those who are nocturnal.
Unlike in mammals, the ciliary muscles in birds eyes attach directly to their cornea without an intermediate ligament like the zonules.[3] This feature allows them a greater range of accommodation. Avian ciliary muscles also are fast muscle fibers, including a high concentration of mitochondria. In some aquatic birds this along with the softness of the avian lens has made it possible for them to compensate for the refractive loss in the cornea while under water. In addition to being able to adjust the shape of their lens with the aid of ciliary muscles, like other vertebrates, some birds also have a set of muscles around the cornea called Crampton's muscles. [3] This feature is not found in aquatic birds, but it is found in species that locate food on the ground, such as the chicken, or birds of prey such as the kestral. Like many mammals, birds have a small blind spot without photo-receptors at the optic disc, where the optic nerve and blood vessels join the eye.
The highly specialized avian eye contains rods, two types of cone receptors - four types of single cones permitting tetrachromatic color vision, and a set of double-cone photoreceptors.[4] Although cones require the presence of more light, they are essential to visual acuity, or "sharpness".
Retina
Avian retinas are similar to most other vertebrates. Birds have relatively large eyes compared to the size of their body which permits a large image to be projected onto the retina. They have a higher density of photoreceptors which is critical in determining visual acuity. The cone cells in birds are extremely important and can account for around 80% of their receptors. Noctural birds like the owl have almost all rods which help with night vision. Light passes through the retina and forms the image on the pigmented epithelium at the back of the eye.(STURKIE) Pigment molecules in the photoreceptors retrieve the image and "transduce" it to become nervous impulses. The fovea has the greatest density of receptors and is located in the center of the retina. Birds have a small blind spot which has no photoreceptors under the optic disc near the optic nerve. Unlike humans, birds do not compensate for the blind spot by filling it in with the surrounding picture the eye processes. Instead, it remains an area with no vision.
Eyelids
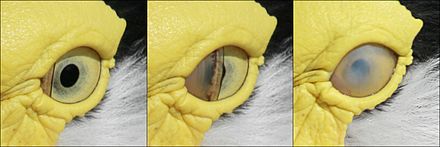
All birds contain three eyelids. The two outer eyelids for birds are not used for blinking. The necessary function of lubricating the eye is instead carried out by the nictitating membrane, or third eyelid. It swipes across the cornea horizontally and is transparent allowing for full time visibility. When not in use, the nictitating membrane stays in the corner of the eye. In humans, this is the visible pink nub in the corner of the eye although it does not have the same functions as the bird’s eye anymore[5]. The nictitating membrane is necessary for survival since losing a fraction of second while flying 60 miles per hour among trees could be fatal. The nictitating membrane also maintains hygiene and cornea protection[5]. It is suspected that some birds may fly with the third eyelid covering the cornea of the eye, which prevents it from drying out during flight, acting like birdy goggles.
Eyelid movement is controlled by 4 striated muscles. The levator palpebrae superioris muscle, which has both skeletal and smooth muscle fibers, elevates the upper eyelid, whereas the depressor palpebrae ventralis muscle, as its name implies, depresses the lower lid. The 2 other muscles, the tensor periorbitae and orbicularis oculi, which may actually be one and the same muscle, encircle the eyelid and are innervated by the mandibular nerve.[6]
The eyes are kept moist by secretions from the Harderian and lachrymal glands. (STURKIE)
Unique Visual Optics
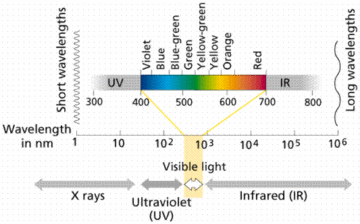
Biologist continue to grapple with the reasons for natures vast diversity of coloration in the natural world. Birds are among the most studied examples of diverse coloration, with some of the most striking array of variation and display in the natural world. They are known for having the most richly endowed visual system of any vertebrate, with high UV-A sensitivity and tetrachromatic vision, able to see wavelength of 300 to 700 nm.[7]
UV Vision

Most birds are tetrachromats or have four types of cone cells; red, green, blue and ultraviolet. These four types of cones are what allows them to process a wider wavelength spectrum than humans. Their spectrum extends to the UV range of 300-400 nm. Although birds are not the only ones who have UV vision (fish, amphibians, reptiles, and insects also have the ability), it does provide them with evolutionary advantages. One of the most important functions is orientation. UV vision gives birds an advantage over trichromats and dichromats in being able to identify shapes and objects which facilitates in travel. Another important function is the increased ability to look for food. UV vision allows birds to more easily spot insects, berries, seeds and mammals[8]. Another function is signaling, especially during mate selection. The UV allows birds to differentiate between the desirable and undesirable candidates for mating.
Pecten Oculi
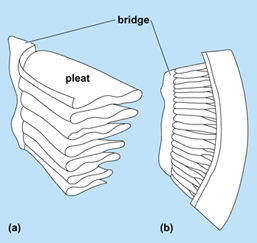
The pecten oculi is a nonsensory, highly vascular, pigmented structure unique to birds and is located in the vitreous chamber. There are three morphological types of pecten oculi in birds; conical, vaned, and pleated[9]. The shapes and number of pleats are different among nocturnal and diurnal birds. Diurnal birds usually have a larger pecten and more folds than nocturnal birds. It is folded and pleated to increase surface area[10]. The pecten oculi moves the blood vessels in the back of the eye resulting in a sharper image[9]. Also, it acts like a central retinal artery in the eye and supplies oxygen to the retina, provides pressure regulation, pH regulation, stabilizes the vitreous, and reduces glare in the eye. It has been suggested that there are 30 more functions the pecten oculi performs[9][10].
The pectin is pigmented with 3 to 30 laminae, assembled into an accordion like structure.
Color Vision
When looking at the visible light spectrum, we can see that different colors correspond to specific wavelengths. But color is not that simple. While humans look at the sky and see blue, birds see it much differently, even though it is the same wavelength hitting each retina. This is a hard concept to realize because we generally think that what we see is the truth, and everyone else must see it the same way because they are looking at the same thing. But color vision is based on how different photoreceptors respond, in comparison to each other, to a specific wavelength and combination of wavelengths. Every type of photoreceptor in every species responds slightly differently to the same wavelength, so the color is perceived differently. Color perception is a very complex process through higher order neuronal processing that has to do with the relationship between photoreceptor, bipolar, ganglion, amacrine and horizontal cells in the retina, as well as the pathways the signals use to travel to the brain. Color is then analyzed in the brain before a specific color is perceived[11].
Types of Photoreceptors
Birds have one of the most sophisticated systems of cone photoreceptors among vertebrates. Their vision is most comparable to reptiles out of all the classes of animals. Most birds are able to see wavelengths over a long range, many times reaching past the visible light range into the ultra-violet light range. It is thought that birds are also able to see polarized light and use magnetoreception to directionally orient themselves which is dependent on wavelengths of ambient light. Many animals are dichromatic, meaning they have two types of cones that each have a different range of wavelengths, while humans and other primates are trichromatic, having 3 types of cones. Because most species of birds have four types of cones, they are tetrachromatic. There is evidence leading to pigeons being pentachromatic with five types of cones, and if this were proven true many people believe that more birds would turn up to be pentachromatic[12]. Therefore, in addition to the rod cells, birds have generally 5, and possibly 6 types of photoreceptors. The 4 types of cones consist of 3 cones similar to ours in the blue, green and red spectrum of visible light, with the 4th cone being either in the violet spectrum or down in the ultra-violet spectrum. In addition to the main types of cones, birds also have double cones. These consist of 2 types of cones known as principal and accessory members. These double cones are two cells attached to each other so they are able to communicate with each other in a very direct way. They use this communication for many different functions that are not all known. There is some thought that they are used for mostly achromatic functions like detecting polarized light and motion detection and that mostly only the single cones are used for detecting color[11]. Others believe these cones do help with color detection. While it is not fully understood what the purpose of these double cones are, it is thought that some of the processing of vision that happens in the brains of humans starts in these photoreceptors of birds[13]. In some species of birds, up to 50% of their cones are double cones[11].
Range of Wavelengths
One thing that makes birds different from most other species is that they have oil droplets in the inner segment of their cones, right on the border between the inner and exterior segments. These oil droplets provide an extra medium for the wavelengths to pass through before reaching visual pigment and activating the cone. The main purpose of the oil droplet is to narrow the range of wavelengths that the cell responds to by absorbing some of the longer wavelengths so they can't pass through. With narrower absorption spectra, the cones have less overlap with each other making it so birds can see a greater number of colors. These oil droplets are colored by different densities of carotenoids to filter out different wavelengths of light. The longer wavelength (red) cones have oil droplets with more carotenoids so they are darker red. The green sensitive cones have yellow oil droplets, blue sensitive cones have oil droplets that are essentially clear or colorless and violet/ultra-violet sensitive cones have transparent oil droplets with no carotenoids [12]. The opsin-like protein in each cone along with the oil droplet determines the maximum absorbance wavelength for that cone type. This maximum is just the peak of the curve of the range of wavelengths that each cone absorbs. In average maximum wavelength absorptions for different species of birds is as follows:
| Receptor Type | Wavelengths |
| LWS (red cones) | 560nm-570nm |
| MWS (green cones) | 500nm-510nm |
| SWS (blue cones) | 430nm-460nm |
| Rods | 500nm-510nm |
| UVS cones (ultra-violet or violet) | violet range – 400nm-420nm ultra-violet range – 360nm-380 |
Photoreceptor distribution on retina
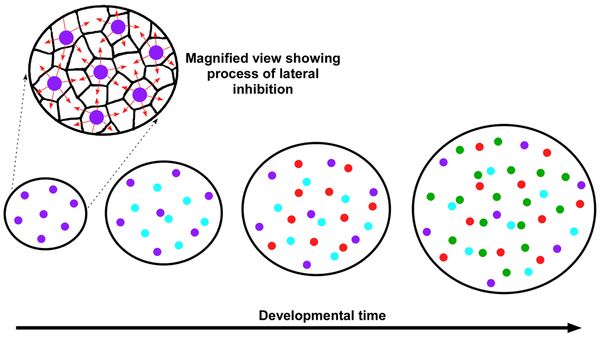
The density of photoreceptors on the retina determines the visual acuity of the animal. Humans have about 200,000 receptors per mm2, while birds have 2 to 5 times this amount. Due to the many reasons that color vision is so important in birds, diurnal birds have a much higher percentage of cones on their retina than rods. About 80%-90% of photoreceptors in diurnal birds are cones. The mosaic of photoreceptors on the retina is not highly studied and is thought to be random in most species. One study using chickens verified with three other species of birds shows that unlike many other species, birds have an extremely high degree of spatial organization of photoreceptors on their retina. This study showed that each type of cone independently tiles the retina with highly regular organization. All five types of cones tile the retina in a similar way but independent of each other. This was determined by comparing the relationship of density with the distance to the nearest neighboring cone of the same type. It was shown that this relationship is very specific and organized. When density increases, the distance between cones of the same type decreases. When graphed, this relationship produces a curve that is very similar in all 5 types of cones. Each type of cone uses a common mechanism to determine the spacing between each cell. It is not fully understood how the cells organize in this way but it is clear that the cone cells have to have two signaling systems in order to create this topographical map. One system needed is to identify neighboring cells in terms of cone type, and the other is a system for measuring the distance between cone cells of the same type. It is thought that cone cells tile the retina using lateral inhibition to ensure the cones appear in the correct pattern [14].
Complexities of Color Perception
Color perception gets even more complex when you take into account plane polarized light and magnetoreception. There is evidence that birds use magnetoreception to orient themselves with the earth so they are able to migrate at night when there is minimal light and their vision worsens. There is also evidence that birds can see plane polarized light, which is light that shifts it's directional orientation as it hits certain medium where small amounts of the light is absorbed. This directional orientation does not change how humans see light, or colors, but for species whose photoreceptors detect these plane polarized changes they may see things much different than we do. Birds detect these differences using the same photoreceptors that detect color, so plane polarized light affects how they perceive colors. This makes it complicated for us to understand how birds see certain colors because there are more things to take into account than just wavelengths of light and types of cones[11].
Effects of Cones on Night Vision
While birds have some of the most highly developed color vision that surpasses that of humans and many other species, there is one disadvantage to this. Only about 20% of the photoreceptors in diurnal birds are rods, which are much more sensitive to light than cones, but cannot detect color. This is a much lower percentage than in humans whose retina contain about 95% rods. This ratio results in birds needing about 5-20 times as much light to see colors as humans do because they have fewer rod cells that can detect much lower amounts of light than cones. Diurnal birds have a much harder time seeing color as twilight approaches than most other species[15].
Nocturnal Birds
Nocturnal Birds have a majority of rods on their retina, so they can see much better at night. Owls have only about 7%-8% cones, which leaves about 90% rods[16].
Motion Detection
Birds have a high flicker fusion rate which creates the ability to see fast moving objects. They can see around 100Hz as continuous movement where humans have the ability to see about 60Hz. Since birds have evolved to move at faster speeds, a high flicker fusion frequency gives birds better visual acuity. They can assimilate detail much faster than a mammal. Birds of prey have a double fovea which allows them to see very slow movements as well. This gives them the ability to keep track of a moving object very easily and helps when migrating. A pigeon has the ability to detect movement as slow as 15 degrees per hour.
Flying/Head Bobbing
To obtain steady images while flying or swaying on a branch, birds hold their head as steady as possible with compensating reflexes. In a variety of species if the eye and the head are consistently maintained in a particular position the entire retina can view the ground in focus at once.[17]Maintaining a steady image is especially important for birds of prey. Head bobbing behavior stabilizes visual fields during body movement, preventing a blur of the retinal imagine. It gives them cues for distance and location of objects. Some birds head bob to achieve motion parallax. For example, pigeons head bob when landing to see depth.
Neuronal Processing
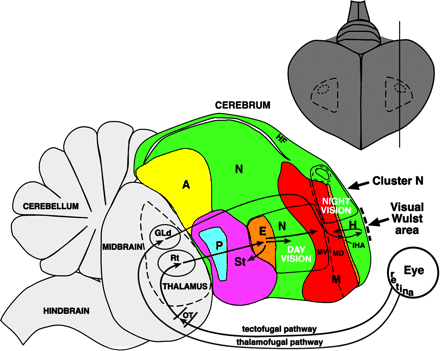
There are two pathways that lead to the eyes - traveling from either the optic tectum, or the dorsal thalamus. The retino-collicular, is the connecting pathway between the retina and the midbrain colliculus, possessing the efferent pathways to the thalamus. From the thalamic center, nuclei project into the telencephalon (also known as the cerebrum). Mammalian vision studies throughout the previous decades have highlighted the dorsal thalamus, striate cortex, and the extrastriate areas for general mammal visual processing. Yet, the avian collothalamic system is thought to be the dominant pathway, and is assumed to be true in lateral-eyed species. [19]
Different patterns of intersection of nerve pathways has lead scientists to believe that binocular vision has developed independently in birds. Most mammals are found to have partial crossover between only the optic nerve fibers from the nasal side project to the contralateral side, while the temporal projects directly to the same side. In birds, most have laterally placed eyes, and experience full crossover to either side of the brain. Predatory birds, like owls, are known for having more forward facing eyes, presumably leading to better binocular vision, and granting them stereoscopic vision. [19]
The avian telencephalon is known to consist of multiple nuclear cell groups. For the visual system, relevant nuclei are ostensibly anatomically separate and spatially distant within the avian telencephalon. These nuclei are actually organized in a more systematic fashion than was previously understood. Thus, there is an extensive columnar continuum involving different telencephalic structures separated by several laminae. Such a column-like organization may exist not only for vision, but other sensory modalities and even non-sensory systems; in particular,the sensory and limbic areas of the avian brain. The expansion of this continuum may be relevant to the enlargement of the telencephalon in some species, such as corvids, that are known for their complex behavior. [20]
General Higher Order Visual Perception
The telencephalon, or the cerebrum, of birds is extremely important to their visio-cognative capacity. Throughout their brain, separate, highly organized channels of the anterior-posterior axis run parallel to the major laminae. Another highly organized section is the efferent projections of the primary telecephalon, which forms substantial columns with both dorsolateral pallium and the lateral basal ganglia. To understand the connections, biologist researched both behavioral and neuronal integration, for a clear picture of the 150 million year evolution of the avian visual system. [2]
Facial Recognition
Recently researches have found evidence that pigeons use parallel channels for processing static (color, texture) and dynamic (motion) images.[2] This segregation extends from the thalamus to the telencephalon. In humans there is considerable interactions between the pathways that code for color and motion but in birds there seems to be less coordination. One result of the segregation of static and dynamic images is that birds cannot recognize global facial features. They are however able to recognize changes in local facial elements such as the eyes and beak. Interestingly, it has been discovered that crows are able to recognize the faces of humans (often researches who have captured them) and remember which humans they have had unpleasant experiences with. [21]
Magnetic Fields
It has been suggested that migratory birds can see magnetic fields to help direct them. When light enters the eyes of birds it excites a molecule called cytochrome shutting it into a state where it can be affected by the earth’s magnetic field. This magnetic field may be perceived as a dark or light spot which lies upon the normal visual field of the bird. It would change when the bird turns it head. A magnetic field is seen as giving a bird a compass so they can migrate properly.[22]
Evolutionary Significance
The role of color vision in mate selection has received much attention from researchers although it is still a highly contentious, multifaceted subject. It has been debated whether or not birds actually use colorful ornamentation during sexual selection. Some researches believe that we are being anthropomorphic when we attribute mate selection to color, insisting that assigning significance to different colors is an entirely human habit. However, many other researchers insist that color is in fact deeply important in mate selection to many species of birds. (Don't worry everyone, I have citations I just haven't learned how to include them yet)
Role of Color in Mate Selection
Color is regarded as a way for birds to convey honest sexual signalling. Red pigment has received much attention because it is often affected by birds' diet. For example in male house finches red pigment is achieved through eating a diet rich in carotenoids. Because birds must also use carotenoids in maintaining individual health, including their vision, there must be a balance struck between concentrating carotenoids in their feathers and using them in the retina, as antioxidants, etc. This trade off is thought to give potential mates an "honest" signal of their health. Males that are able to afford brightly colored feathers are likely to also be good foragers and to be in good health. Additionally, the concentration of carotenoids in both sexes of house finches peaks in November during their period of prebreeding mate selection. It is hypothesized that female house finches are better able to perceive male's coloration during this time because they also have heightened levels of carotenoids.
Color also plays a role in the selection of mates by males. Specifically in hole nesting birds, because of poor light quality in the nest, females with pale-blue eyes are more visible. It is believed that males perceive this to mean the female is healthy and fertile. This is one male-provisioning selections among the birds. [23] Scientists have come to 3 hypothesis for the variations in egg color as a whole. First, it is thought that birds being able to see in the ultraviolet light spectrum causes an interaction between various properties of avian vision and the light environment in the nest. Second, the blueish-green tint of eggs is believed to be because of sexually select signals to males regarding female genetic quality. And finally, the instance of spots on eggshells is thought the be of structural benefit to eggs, compensating for thinning due to calcium issues.[23]
Influence of Ultra Violet Light Perception
Species of birds that have been found to have ultra violet vision are much more likely to have ultra violet coloring than species that are unable to perceive UV wavelengths. Nocturnal birds, for instance need to perceive differences in light and dark rather than in color and do not perceive ultra violet wavelengths, nor do they exhibit UV pigments in their feathers. In contrast, birds with bright colors in the visible spectrum frequently also have ultra violet pigments that human and other trichormatic species cannot perceive.
While colors in the ultra violet range in feathers or in bird's skin can be a positive indication of health, UV pigment can also be an indication of an undesirable mate. For example, in male red grouse the appearance of ultra violet pigments in their combs and the skin beneath their wings signals that they are not the fittest mate. This is because UV pigment is located in their dermis, while red carotenoids color their epidermis. Individuals that exhibit UV coloring are lacking in carotenoids which implies that they are unhealthy or poor foragers. In this case females would select red grouse with red colored combs but not select individuals with prominent ultra violet coloring.
Sexual Dichromatism
Sexual dichromatism, the common phenomena of male and female birds within the same species exhibiting different ornamentation, is one example of how tetrachromacy has influenced the mating habits of birds along with their physical appearance. Sexual dichromacy may occur when one sex is selected for either more ornamentation or less ornamentation [24]. Additionally, ultra violet pigment undetectable to mammals may be used by birds to distinguish between sexes. This occurs in species like the blue tit and starlings, where females are attracted to males with stronger ultra violet reflectance.
Odds N' Ends
Most birds' eyes are located on the sides of their heads, with one fovea. Hawks, swallows, and hummingbirds, have a second fovea called the temporal fovea that translates images from the front of the bird, specific to those who "feed on the wing". [1] The bird species with the greatest field of vision is the American woodcock Its eyes are positioned exactly opposite of each other, allowing it an horizontal visual field of 360° and a vertical field of vision of 180°, which enables it to quickly scan its surrounding area without moving its eyes or head.
A researcher named Watanabe was able to train pigeons to differentiate between paintings of Monet (Impressionism) and Picasso (Cubism). He went so much as to further test their knowledge - testing the pigeons knowledge of paintings not used in the training and continued to be able to generalize the classification rules they had learned and correctly identify the two styles of painting. (WATANABE) This information is thought to depict avian ability to gain visual concept of objects throughout their life. Evolutionarily it is important for hunting, migrating and surviving from predators.
Possible Test Questions
Question: Why is it important for birds to have a transparent eyelid for blinking?
Question: When talking about vision, what is another name for the cerebrum?
Question: What role do the oil droplets in the cones of birds play and what are some advantages they provide?
References
- ↑ 1.0 1.1 1.2 Eldon, G. (2011). Avian Vision. Birder's World, 25(1), 44–47, <a href="http://search.ebscohost.com/login.aspx?direct=true&db=fzh&AN=57232317&site=ehost-live">Avian vision.</a>.
- ↑ 2.0 2.1 2.2 Shimizu, T., Patton, T. B., Husband, S. A. (2010). “Avian Visual Behavior and the Organization of the Telencephelon.” Brain, Behavior & Evolution, 75(3), 204-218. Print
- ↑ 3.0 3.1 Martin, G. R. (2009). What is binocular vision for? A birds’ eye view. Journal of Vision, 9(11):14, 1–19, http://journalofvision.org/9/11/14/.
- ↑ Knott, B. Berg, M. L., Morgan, E. R., Buchanan, K. L., Bowmaker, J. K., Bennett, A. T., 2010, Avian Retina Oil Droplets: Dietary manipulation of color vision, Proceedings of the Royal Society of Biological Sciences Series B. 277, 953-962, print.
- ↑ 5.0 5.1 E. Philippe Stibbe. “A Comparitive Study of the Nictitating Membrane of Birds and Mammals.” 160-164.
- ↑ Michael P. Jones, Kenneth E. Pierce Jr, Daniel Ward, Avian Vision: A Review of Form and Function with Special Consideration to Birds of Prey, Journal of Exotic Pet Medicine, Volume 16, Issue 2, April 2007, Pages 69-87, ISSN 1557-5063, 10.1053/j.jepm.2007.03.012.
- ↑ Lina, O. (2009). Avian Color Vision: Effects of Variation in Receptor Sensitivity and Noise Data on Model Predictions as Compared to Behavioral Results, Vision Research, 49(15), 1939-1947, print.
- ↑ Stuart C. Church, Annabels L. Merrison and Toby M.M. Chamberlain. “Avian Ultraviolet Vision and Frequency-Dependent Seed Preferences.” The Journal of Experimental Biology (2001) 2491–2498.
- ↑ 9.0 9.1 9.2 P. C. O. C. Jezler, M. B. P.Braga, E. Perlmann, R. Squarzoni, M. I. Borella, P. S. M. Barros, L.Milanelo, and A. Antunes. “Histological Analysis of Eyeballs of the Striped Owl.” Formatex (2010): 1047-1054. http://www.formatex.info/microscopy4/1047-1054.pdf
- ↑ 10.0 10.1 Vincent Brach. “Fundamental Significance of Pecten: A Review” (1996): 321-326.
- ↑ 11.0 11.1 11.2 11.3 Hill, Geoffrey E., and Kevin J. McGraw. Bird Coloration. Cambridge, MA: Harvard UP, 2006. Print.
- ↑ 12.0 12.1 12.2 Bowmaker, J.k., L.a. Heath, S.e. Wilkie, and D.m. Hunt. "Visual Pigments and Oil Droplets from Six Classes of Photoreceptor in the Retinas of Birds." Vision Research 37.16 (1997): 2183-194. Print.
- ↑ "Color Vision in Birds." Causes of Color. Web Exhibits. Web. 01 Dec. 2011. <http://www.webexhibits.org/causesofcolor/17B.html>.
- ↑ 14.0 14.1 Kram, Yoseph A., Stephanie Mantey, and Joseph C. Corbo. "Avian Cone Photoreceptors Tile the Retina as Five Independent, Self-Organizing Mosaics." Ed. Eric Warrant. PLoS ONE 5.2 (2010). Print.
- ↑ The Swedish Research Council. "Birds lose color vision in twilight." ScienceDaily, 11 Nov. 2009. Web. 1 Dec. 2011.
- ↑ Jacobs, Gerald H., Michael Crognale, and John Fenwick. "CONE PIGMENT OF THE GREAT HORNED OWL." The Condor (1987). Print.
- ↑ "The Eye: A Survey of Human Vision." Google Books. EM Publications. Web. 22 Nov. 2011.fckLR
- ↑ Nervous System: Brain and Special Senses II <http://people.eku.edu/ritchisong/birdbrain2.html>
- ↑ 19.0 19.1 "V. Taking Flight: Post-Retinal Processing" Copyright 2004 <http://www.pigeon.psy.tufts.edu/avc/husband/avc5vpth.htm>
- ↑ Toru Shimizu,a* Tadd B. Patton,a and Scott A. Husbandb "Avian Visual Behavior and the Organization of the Telencephalon." Department of Psychology, University of South Florida, Tampa, Fla., USABrain Behav Evol. 2010 August; 75(3): 204–217. Published online 2010 August 20.<http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2977968/>
- ↑ Berry, B. (2009). Inter-species and Intra-species Appearance Bias: Social Learning, Interactionism, and Other Sociological Interpretations. Conference Papers -- American Sociological Association, 1.
- ↑ Bird Vision: Bird Anatomy. General LLC, 2010. Print.
- ↑ 23.0 23.1 Cherry, M. I., Goslerm A. G., 2010, Avian Eggshell Coloration: New Perspectives on Adaptive Explanations, Biological Journal of The Linnean Society, 100(4). 753-762. Print.
- ↑ Avian Sexual Dichromatism in Relation to Phylogeny and Ecology. Alexander V. Badyaev and Geoffrey E. Hill. Annual Review of Ecology, Evolution, and Systematics , Vol. 34, (2003), pp. 27-49
